Ciclo del ácido cítrico
El ciclo del ácido cítrico — también conocido como el ciclo del ácido tricarboxílico (Ciclo TCA), o el Ciclo de Krebs,[1][2] — es una serie de reacciones químicas utilizado por todos organismos aerobios para generar energía a través de la oxidación de acetato de deriva hidratos de carbono, grasas y proteínas en dióxido de carbono y energía química en forma de trifosfato de adenosina (ATP). Además, proporciona el ciclo precursores de ciertos aminoácidos, así como la agente reductor NADH se utiliza en numerosas otras reacciones bioquímicas. Su importancia central en muchas vías bioquímicas sugiere que era uno de los primeros componentes establecidos de celulares metabolismo y puede tener su origen abiogenically.[3]
Se deriva el nombre de este camino metabólico ácido cítrico (un tipo de ácido tricarboxílico) que se consume y luego regenerada por esta secuencia de reacciones para completar el ciclo. Además, el ciclo consume (en forma de acetato Acetil-CoA) y agua, reduce NAD+ NADH, y produce dióxido de carbono como un subproducto inútil. El NADH generado por el ciclo TCA es alimentado en el fosforilación oxidativa vía (transporte de electrones). El resultado neto de estas dos vías estrechamente vinculados es la oxidación de nutrientes para producir energía química utilizable en forma de ATP.
En eucariotas las células, se produce el ciclo del ácido cítrico en la matriz de la mitocondria. En células procarióticas, como las bacterias que carecen de mitocondrias, la secuencia de la reacción del TCA se realiza en el citosol con el gradiente de protones para la producción de ATP en (superficie de la célulamembrana plasmática) en lugar de la membrana interna de la mitocondria.
Contenido
- 1 Descubrimiento
- 2 Evolución
- 3 Resumen
- 4 Pasos
- 5 Productos
- 6 Eficiencia
- 7 Reglamento
- 8 Principales vías metabólicas convergiendo sobre el ciclo del TCA
- 9 Mapa interactivo vía
- 10 Véase también
- 11 Referencias
- 12 Enlaces externos
Descubrimiento
Varios de los componentes y las reacciones del ciclo de ácido cítrico se establecieron en la década de 1930 por la investigación de la premio Nobel Albert Szent-Györgyi, para la cual recibió el Premio Nobel en 1937 por sus descubrimientos relativos a ácido fumárico, un componente clave del ciclo.[4] El ciclo del ácido cítrico se fue finalmente identificado en 1937 por Hans Adolf Krebs mientras que en el Universidad de Sheffield, para lo cual recibió el Premio Nobel de Fisiología o medicina en 1953.[5]
Evolución
Componentes del ciclo TCA fueron derivados bacterias anaerobias, y el ciclo TCA se puede haber evolucionado más de una vez.[6] Teóricamente existen varias alternativas para el ciclo del TCA; Sin embargo, el ciclo TCA parece ser la más eficiente. Si varias alternativas TCA habían evolucionado independientemente, todos ellos parecen tener convergentes para el ciclo TCA.[7][8]
Resumen
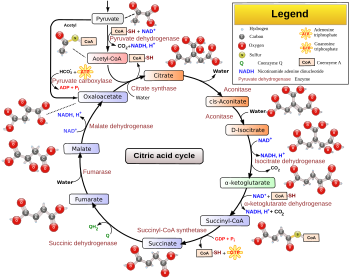
El ciclo del ácido cítrico es una vía metabólica clave que unifica a hidratos de carbono, grasa y metabolismo proteico. Las reacciones del ciclo se realizan por 8 enzimas que oxidan completamente Acetil-CoA en las dos moléculas de dióxido de carbono. A través de catabolismo de azúcares, grasas y proteínas, se produce un acetato 2 carbones producto orgánico en forma de acetil-CoA que entra en el ciclo del ácido cítrico. Las reacciones del ciclo también convierte tres equivalentes de Nicotinamida adenina dinucleótido (NAD+) en tres equivalentes de NAD reducido+ (NADH), un equivalente de FLAVINA-adenina-dinucleótido(FAD) en un equivalente de FADH2y un equivalente de difosfato de guanosina (PIB) e inorgánicos fosfato (P)i) en un equivalente de guanosín trifosfato (GTP). El NADH y FADH2 generado por el ciclo del ácido cítrico a su vez son utilizados por el fosforilación oxidativa camino a generar a ricos en energía trifosfato de adenosina (ATP).
Una de las fuentes primarias de acetil-CoA proviene de la descomposición de azúcares por glucólisis que ceder piruvato a su vez es descarboxilada por la enzima Piruvato deshidrogenasa generación de acetil-CoA según el siguiente esquema de reacción:
- CH3C(=O) C(=O) O– (piruvato) + HSCoA + NAD+ → CH3C(=O) SCoA (acetil-CoA) + NADH + CO2
El producto de esta reacción, acetil-CoA, es el punto de partida para el ciclo del ácido cítrico. Acetil-CoA también puede obtenerse de la oxidación de ácidos grasos. A continuación es un resumen esquemático del ciclo:
- El ácido cítrico comienza el ciclo con la transferencia de un dos-carbono acetil Grupo de Acetil-CoA para el aceptador de cuatro carbonos compuesto (oxaloacetato) para formar un seis-carbono compuesto (citrato).
- El citrato de entonces pasa a través de una serie de transformaciones químicas, perdiendo dos carboxilo grupos como CO2. Los electrodos de carbono perdieron como CO2 se originan en lo que fue oxaloacetato, no directamente de acetil-CoA. Los carbonos donados por acetil-CoA a formar parte de la columna vertebral carbono oxaloacetato después de la primera vuelta el ciclo del ácido cítrico. Pérdida de los carbonos donados por acetyl-CoA como CO2 requiere varias vueltas el ciclo del ácido cítrico. Sin embargo, debido al papel del ciclo del ácido cítrico en anabolismo, podrían no estar perdidos, desde muchos ciclo TCA intermedios se utilizan también como precursores para la biosíntesis de otras moléculas.[9]
- La mayoría de la energía a su disposición por los pasos del ciclo oxidativos se transmite como rico en energía electrones a NAD+, formando NADH. Para cada grupo acetilo que entra en el ciclo del ácido cítrico, se producen tres moléculas de NADH.
- Los electrones se transfieren también al aceptor de electrones Q, formando QH2.
- Al final de cada ciclo, el oxaloacetato cuatro emisiones de carbono ha sido regenerado, y el ciclo continúa.
Pasos
Dos carbono los átomos son oxidado Para CO2, la energía de ser trasladado a otros procesos metabólicos por estas reacciones GTP (o ATP) y como los electrones en NADH y QH2. El NADH generado en el ciclo del TCA puede donar más tarde sus electrones en fosforilación oxidativa para impulsar la síntesis de ATP; FADH2 se une covalentemente a Succinato deshidrogenasa, una enzima que funciona tanto en el ciclo del TCA y el mitocondrial cadena de transporte de electrones en la fosforilación oxidativa. FADH2, por lo tanto, facilita la transferencia de electrones para coenzima Q, ¿cuál es el aceptor de electrones final de la reacción catalizada por la oxidorreductasa succinato: Ubiquinona complejo, también actúa como un intermediario en la cadena de transporte de electrones.[10]
El ciclo del ácido cítrico se suministra continuamente con nueva de carbono en forma de acetil-CoA, entrando en el paso 1 por debajo.[11]
| Sustratos | Productos | Enzima | Tipo de reacción | Comentario | |
|---|---|---|---|---|---|
| 1 | Oxaloacetato + Acetil CoA + H2O |
Citrato + CoA-SH |
Citrato sintasa | Condensación aldólica | irreversible, se extiende el oxaloacetato 4C a una molécula de 6C |
| 2 | Citrato | CIS-Aconitato + H2O |
Aconitasa | Deshidratación | isomerización reversible |
| 3 | CIS-Aconitato + H2O |
Isocitrato | Hidratación | ||
| 4 | Isocitrato + NAD+ |
Oxalosuccinate + NADH + H + |
Isocitrato deshidrogenasa | Oxidación | genera NADH (equivalente a 2,5 ATP) |
| 5 | Oxalosuccinate | Α-Cetoglutarato + CO2 |
Descarboxilación | etapa tarifa-limitación, irreversible, genera una molécula de 5C |
|
| 6 | Α-Cetoglutarato + NAD+ + CoA-SH |
Succinil-CoA + NADH + H+ + CO2 |
Α-Cetoglutarato deshidrogenasa | Oxidativo descarboxilación |
etapa irreversible, genera NADH (equivalente a 2,5 ATP), regenera la cadena 4C (CoA excluido) |
| 7 | Succinil-CoA + PIB + Pi |
Succinato + CoA-SH + GTP |
Sintetasa Succinil-CoA | nivel de sustrato fosforilación | o ADP→ATP en lugar de GDP→GTP,[10] genera 1 ATP o equivalente Reacción de condensación de PIB + Pi y hidrólisis de Succinil-CoA involucrar a la H2O necesarios para la ecuación balanceada. |
| 8 | Succinato + Ubiquinona (Q) |
Fumarato + Ubiquinol (QH2) |
Succinato deshidrogenasa | Oxidación | usos FAD como un grupo prostético (FAD→FADH2 en el primer paso de la reacción) de la enzima,[10] genera el equivalente a 1,5 ATP |
| 9 | Fumarato + H2O |
L-Malato | Fumarasa | Hidratación | |
| 10 | L-Malato + NAD+ |
Oxaloacetato + NADH + H+ |
Malato deshidrogenasa | Oxidación | reversible (de hecho, el equilibrio favorece malato), genera NADH (equivalente a 2,5 ATP) |
Las mitocondrias en los animales, incluidos los seres humanos, poseen dos síntesis de Succinil-CoA: una que produce GTP del PBI y otra que produce ATP de ADP.[12] Las plantas tienen el tipo que produce ATP (formación de ADP sintetasa de Succinil-CoA).[11] Varias de las enzimas en el ciclo pueden asociar libremente en un multienzyme complejo de la proteína dentro de la matriz mitocondrial.[13]
El GTP que está formado por la formación del PIB sintetasa Succinil-CoA puede ser utilizado por quinasa nucleósido difosfato para formar ATP (la reacción catalizada es GTP + ADP → PIB + ATP).[10]
Productos
Los productos de la primera vuelta del ciclo son: uno GTP (o ATP), tres NADH, una QH2CO dos2.
Porque dos acetil-CoA moléculas son producidos por cada uno glucosa molécula, dos ciclos son necesarios por la molécula de glucosa. Por lo tanto, al final de dos ciclos, los productos son: dos GTP, 6 NADH, dos QH2y cuatro CO2
| Descripción | Reactantes | Productos |
| Es la suma de todas las reacciones en el ciclo del ácido cítrico: | Acetil-CoA + 3 NAD+ + Q + PIB + Pi + 2 H2O | → CoA-SH + 3 NADH + H 3+ + QH2 + GTP + 2 CO2 |
| Combinando las reacciones que ocurren durante el oxidación del piruvato con los que se producen durante el ciclo del ácido cítrico, se obtiene la reacción de oxidación del piruvato generales siguientes: | Ion piruvato + 4 NAD+ + Q + PIB + Pi + 2 H2O | → 4 NADH + H 4+ + QH2 + GTP + 3 CO2 |
| Combinando la reacción anterior con los que ocurren en el curso de glucólisis, el siguiente en general reacción de oxidación de la glucosa (excluyendo las reacciones en la cadena respiratoria) se obtiene: | Glucosa + 10 NAD+ + 2 Q + 2 ADP + 2 PIB + 4Pi + 2 H2O | → 10 NADH + H 10+ + 2 QH2 + 2 ATP + 2 GTP + 6 CO2 |
Las reacciones anteriores son equilibradas si Pi representa el H2PO4- ion, ADP y PIB el ADP2- y el PIB2- iones, respectivamente y el ATP y el GTP el ATP3- y GTP3- iones, respectivamente.
El número total de ATP obtenido después de la oxidación completa de una glucosa en la glicolisis, ciclo del ácido cítrico, y fosforilación oxidativa se estima que entre 30 y 38.[14]
Eficiencia
El rendimiento máximo teórico de ATP por oxidación de una molécula de glucosa en la glicolisis, ciclo del ácido cítrico, y fosforilación oxidativa 38 (asumiendo 3 molares equivalentes de ATP por equivalentes NADH y 2 ATP por FADH2). En eucariotas, se generan dos equivalentes de NADH en glucólisis, que lleva a cabo en el citoplasma. Transporte de estos dos equivalentes en la mitocondria consume dos equivalentes de ATP, reduciendo así la producción neta de ATP a 36. Además, las ineficiencias en fosforilación oxidativa debido a la pérdida de protones a través de la membrana mitocondrial y deslizamiento de la ATP sintasabomba /Proton comúnmente reduce la producción de ATP por NADH y FADH2 a menos que el máximo rendimiento teórico.[14] Los rendimientos observados son, por lo tanto, más cerca a ~2.5 ATP por NADH y ~1.5 ATP por FADH2, reduciendo aún más la producción total neta de ATP a aproximadamente 30.[15] Una evaluación de la producción total de ATP con cocientes del protón-a-ATP recién revisadas proporciona una estimación de 29,85 ATP por molécula de glucosa.[16]
Reglamento
La regulación del ciclo TCA está determinada en gran medida por la inhibición del producto y la disponibilidad de sustrato. Si el ciclo se les permitió correr desenfrenado, grandes cantidades de energía metabólica podrían ser desperdiciados en la superproducción de la coenzima reducida como NADH y ATP. El sustrato eventual principal del ciclo es que consigue convertir en ATP ADP. Una cantidad reducida de ADP causa acumulación de precursores NADH que a su vez puede inhibir un número de enzimas. NADH, un producto de Deshidrogenasas todos en el ciclo del TCA con excepción de Succinato deshidrogenasa, inhibe Piruvato deshidrogenasa, isocitrato deshidrogenasa, Α-Cetoglutarato deshidrogenasay también citrato sintasa. Acetil-CoA inhibe Piruvato deshidrogenasa, mientras que Succinil-CoA inhibe la alfa-cetoglutarato deshidrogenasa y citrato sintasa. Cuando está probado in vitro con enzimas TCA, inhibe la ATP citrato sintasa y Α-Cetoglutarato deshidrogenasa; Sin embargo, los niveles de ATP no cambiar más del 10% en vivo entre el descanso y el ejercicio vigoroso. No hay ninguna conocida alostérica mecanismo que puede explicar grande cambios en la tarifa de la reacción de un alostérica efectoras cuya concentración cambia menos del 10%.[17]
Calcio se utiliza como regulador. Los niveles de calcio de la matriz mitocondrial pueden llegar a las decenas de micromolar niveles durante la activación celular.[18] Activa fosfatasa piruvato deshidrogenasa que a su vez activa la complejo piruvato deshidrogenasa. Calcio también activa isocitrato deshidrogenasa y Α-Cetoglutarato deshidrogenasa.[19] Esto aumenta la velocidad de reacción de muchos de los pasos en el ciclo y por lo tanto aumenta el flujo a lo largo de la vía.
Citrato es utilizado para la inhibición de la regeneración, como inhibe Fosfofructoquinasa, una enzima implicada en glucólisis esa formación de catalizador de fructosa 1, 6-bifosfato, un precursor del piruvato. Esto evita una constante alta tasa de flujo cuando hay una acumulación de citrato y una disminución en el sustrato de la enzima.
Trabajo reciente ha demostrado un vínculo importante entre intermedios del ciclo de ácido cítrico y la regulación de factores inducible por la hipoxia (HIF). HIF juega un papel en la regulación de la homeostasis del oxígeno y es un factor de transcripción objetivos angiogénesis, remodelado vascular, utilización de la glucosa, transporte del hierro y la apoptosis. HIF es consititutively sintetizado e hidroxilación de al menos uno de dos residuos de prolina crítica interviene en su interacción con el von Hippel Lindau E3 ubiquitina ligasa complejo, que los objetivos para su rápida degradación. Esta reacción es catalizada por Prolyl 4-hidroxilasas. Fumarato y succinato han sido identificados como inhibidores potentes de la prolil hidroxilasas, llevando así a la estabilización de HIF.[20]
Principales vías metabólicas convergiendo sobre el ciclo del TCA
Varios catabólica caminos convergen sobre el ciclo del TCA. Se denominan reacciones que forman intermedios del ciclo TCA para reponerlos (especialmente durante la escasez de los productos intermedios) reacciones anapleróticas.
El ciclo del ácido cítrico es el tercer paso en catabolismo de hidratos de carbono (la desagregación de los azúcares). Glucólisis descompone la glucosa (una seis-carbono-molécula) en piruvato (una molécula de tres carbonos). En eucariotas, piruvato se traslada a la mitocondria. Se convierte en acetil-CoA por descarboxilación y entra en el ciclo del ácido cítrico.
En catabolismo proteico, proteínas están desglosadas por proteasas en sus aminoácidos constituyentes. Las espinas dorsales de carbono de algunos de estos los aminoácidos puede convertirse en una fuente de energía por ser convertidos en acetil-CoA y entrando en el ciclo del ácido cítrico.
En catabolismo de grasa, triglicéridos son hidrolizado para romperlas ácidos grasos y glicerol. En el hígado el glicerol puede ser convertido en glucosa vía Dihidroxiacetona fosfato y gliceraldehído-3-fosfato por medio de gluconeogénesis. En muchos tejidos, especialmente el tejido cardíaco, los ácidos grasos se descomponen a través de un proceso conocidos como beta oxidación, que se traduce en acetil-CoA, que puede ser utilizado en el ciclo del ácido cítrico. Beta oxidación de ácidos grasos con un número impar de puentes metileno produce propionil CoA, que luego es convertida en Succinil-CoA y alimentados en el ciclo del ácido cítrico.[21]
La energía total obtenida de la ruptura completa de una molécula de glucosa por glucólisis, ciclo del ácido cítrico y la fosforilación oxidativa equivale a cerca de 30 moléculas de ATP, en eucariotas. El ciclo del ácido cítrico se llama un Anfibolismo vía porque participa en ambos catabolismo y anabolismo.
Mapa interactivo vía
Haga clic en los genes, proteínas y metabolitos abajo para enlazar con los artículos respectivos. [§ 1]

- ^ El mapa interactivo vía puede editarse en WikiPathways: "TCACycle_WP78".
Véase también
- Ciclo de Calvin
- Ciclo del glioxilato
- Ciclo de Krebs inverso (reductiva)
Referencias
- ^ Lowenstein JM (1969). Métodos en enzimología, volumen 13: ciclo del ácido cítrico. Boston: Prensa académica. ISBN0-12-181870-5.
- ^ Krebs, Weitzman PDJ (1987). Ciclo del ácido cítrico de Krebs: medio siglo y todavía da vuelta. Londres: Sociedad bioquímica. ISBN0-904498-22-0.
- ^ Lane, Nick (2009). Vida ascendente: Los diez grandes inventos de la evolución. Nueva York: W.W. Norton & Co. ISBN0-393-06596-0.
- ^ "El Premio Nobel de Fisiología o medicina 1937". La Fundación Nobel. 26 / 10 / 2011 obtenido.
- ^ "El Premio Nobel de Fisiología o medicina 1953". La Fundación Nobel. 26 / 10 / 2011 obtenido.
- ^ Gest H (1987). "Raíces evolutivas del ciclo de ácido cítrico en procariotas". Biochem. Soc Symp. 54:: 3 – 16. PMID3332996.
- ^ Meléndez-Hevia E, Waddell TG, Cascante M (septiembre de 1996). "El enigma del ciclo del ácido cítrico Krebs: ensamblar las piezas de reacciones químicamente factibles y oportunismo en el diseño de las rutas metabólicas durante la evolución". J. análizar Evol. 43 (3): 293-303. Doi:10.1007/BF02338838. PMID8703096.
- ^ O Ebenhöh, Heinrich R (enero de 2001). "Optimización evolutiva de las rutas metabólicas. Reconstrucción teórica de la estequiometría de ATP y NADH produciendo sistemas". Toro. Matemáticas. Biol. 63 (1): 21 – 55. Doi:10.1006/BULM.2000.0197. PMID11146883.
- ^ Wolfe RR, Jahoor F (febrero de 1990). "Recuperación de CO2 marcado durante la infusión de acetato C-2-labeled C-1-vs: implicaciones para los estudios tracer de oxidación del substrato". AM j Clin. NUTR. 51 (2): 248 – 52. PMID2106256.
- ^ a b c d Stryer L, Berg J, Tymoczko JL (2002). Bioquímica. San Francisco: W.H. Freeman. ISBN0-7167-4684-0.
- ^ a b Jones RC, Buchanan BB, Gruissem W (2000). Bioquímica y biología molecular de plantas (1ª ed.). Rockville, Md: Sociedad Americana de fisiólogos de plantas. ISBN0-943088-39-9.
- ^ Milavetz BI, Johnson JD, Mehus JG, Tews K Lambeth (octubre de 1998). "Evidencia genética para la expresión del ATP y GTP específicos Succinil-CoA sintetasas en eucariotas pluricelulares". J Biol Chem. 273 (42): 27580 – 6. Doi:10.1074/JBC.273.42.27580. PMID9765291.
- ^ Barnes SJ, Weitzman PD (junio de 1986). "Organización de las enzimas del ciclo del ácido cítrico en un clúster multienzimático". FEBS Lett. 201 (2): 267 – 70. Doi:10.1016/0014-5793 (86) 80621-4. PMID3086126.
- ^ a b Porter RK, marca MD (septiembre de 1995). "La conductancia del protón mitocondriales y H + / cociente O son independientes de la tarifa de transporte de electrones en los hepatocitos aislados". Biochem. J. 310 (2): 379 – 82. PMC1135905. PMID7654171.
- ^ Stryer L, Berg JM, Tymoczko JL (2002). "Sección 18.6: la regulación de la respiración celular se rige principalmente por la necesidad de ATP". Bioquímica. San Francisco: W.H. Freeman. ISBN0-7167-4684-0.
- ^ Rico PR (diciembre de 2003). "La maquinaria molecular de la cadena respiratoria de Keilin". Biochem. Soc Trans. 31 (Pt 6): 1095 – 105. Doi:10.1042/BST0311095. PMID14641005.
- ^ Voet d., Voet JG (2004) Bioquímica (3ª ed.). Nueva York: John Wiley & Sons, Inc. p. 615.
- ^ Ivannikov, M. et al (2013). "Ca libre mitocondrial2+ Niveles y sus efectos sobre el metabolismo de energía en los terminales nerviosos del Motor de Drosophila". Biophys. J. 104 (11): 2353 – 2361. Doi:10.1016/j.bpj.2013.03.064. PMC3672877. PMID23746507.
- ^ Denton RM, PJ Randle, puentes BJ, Cooper RH, AL Kerbey, Pask HT, Severson DL, Stansbie D, S Whitehouse (octubre de 1975). "Regulación de mamíferos piruvato deshidrogenasa". Análizar la célula. Biochem. 9 (1): 27 – 53. Doi:10.1007/BF01731731. PMID171557.
- ^ Koivunen P, Hirsilä M, Remes AM, Hassinen IE, Kivirikko KI, Myllyharju J (febrero de 2007). "La inhibición del factor inducible por la hipoxia (HIF) hidroxilasas por intermediarios del ciclo del ácido cítrico: posibles vínculos entre el metabolismo de la célula y la estabilización de HIF". J Biol Chem. 282 (7): 4524-32. Doi:10.1074/jbc.M610415200. PMID17182618.
- ^ Halarnkar PP, GJ Blomquist (1989). "Aspectos comparativos del metabolismo propionato". Comp Biochem. Physiol, B 92 (2): 227 – 31. Doi:10.1016/0305-0491 (89) 90270-8. PMID2647392.
Enlaces externos
- Una animación del ciclo del ácido cítrico en Smith College
- Variantes del ciclo del ácido cítrico en MetaCyc
- Las vías conexión al ciclo del ácido cítrico en Enciclopedia de Kyoto de Genes y genomas
- Introducción a Khan Academy
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ciclo del ácido cítrico Vía metabólica | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxaloacetato | Malato | Fumarato | Succinato | Succinil-CoA | ||||||||||||||||||
| Acetil-CoA | NADH + H+ | NAD+ | H2O | FADH2 | FAD | CoA + ATP(GTP) | Pi + ADP(GDP) | |||||||||||||||
| + | H2O | NADH + H+ + CO2 | ||||||||||||||||||||
| CoA | NAD+ | |||||||||||||||||||||
| H2O | H2O | NAD+ | NAD (P) H + H+ | CO2 | ||||||||||||||||||
| Citrato | CIS-Aconitato | Isocitrato | Oxalosuccinate | Α-Cetoglutarato | ||||||||||||||||||
|
||||||||||||||||||||||
|
||||||||||||

|
||||||||||||||||||||||||||||||||||||||||||||